Archives
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2018-07
-
br Material and methods br Results br Discussion Cysticerci
2020-01-08
Material and methods Results Discussion Cysticerci located in host tissues are exposed to several effector mechanisms of the immune system (Flisser et al., 1986; Sciutto et al., 2000; Amit et al., 2011; Singh et al., 2013). In addition, the extensive uptake of host proteins by cysticerci ha
-
The role of ETB clearing
2020-01-08
The role of ETB clearing receptors has been studied in detail in endothelial cell–specific ETB knock-out mice. In these animals, clearance of an intravenous palovarotene of labeled ET-1 was reduced significantly compared with wild-type controls. Importantly, functioning ETB were retained on all oth
-
Considering these reports and with the aim of
2020-01-07
Considering these reports, and with the aim of further investigating the mechanism by which the cAMP-Epac/PKA pathway activates eNOS, we have performed imaging experiments evaluating the effect of drugs that increase cAMP or modify its signalling pathways (PKA or Epac activators and inhibitors) on b
-
zd 14 br Significance Ubl post translational modifications a
2020-01-07
Significance Ubl post-translational modifications are potential targets for developing novel therapeutics for life-threatening diseases, such as c-Myc- and KRas-driven cancers that lack targeted therapy. However, only a few drugs are available to target these modifications. This lack of drugs hig
-
Several large scale clinical studies have set out with
2020-01-06
Several large-scale clinical studies have set out with the common goal to further understand the cognitive impact of menopause and hormone therapies taken by women (Gleason et al., 2015, Greendale et al., 2011, Karlamangla et al., 2017, Rapp et al., 2003, Shumaker et al., 2003). Only one human study
-
P H is a nonheme iron II dioxygenase that
2020-01-06
P4H is a nonheme iron(II) dioxygenase that uses O2 and α-ketoglutarate as co-substrates (Fig. 1A). The three-dimensional structure of mammalian P4H is unknown. We reasoned that an electrophilic analog of α-ketoglutarate could serve as an irreversible inhibitor of the enzyme and, hence, a useful prob
-
A recent report has shown that C elegans homozygous
2020-01-06
A recent report has shown that C. elegans homozygous for a point mutation within the HORMA domain of HTP-1 (M127K), which disrupts its association with chromosome axes, displayed an extended zone of PLK-2 localization to PCs in response to asynapsis (Silva et al., 2014), which, as we show here, is a
-
Currently much effort has been made to design
2020-01-06
Currently, much effort has been made to design and synthesize ThDP analogs (such as ThTDP, ThTTDP, and triazole-ThDP in Fig. 2)8, 9, 10, 11, 12 as PDHc E1 inhibitors. These ThDP analog inhibitors, such as ThTDP and ThTTDP, can block the ThDP binding site, and exhibit significantly stronger binding a
-
br These component origins have been
2020-01-06
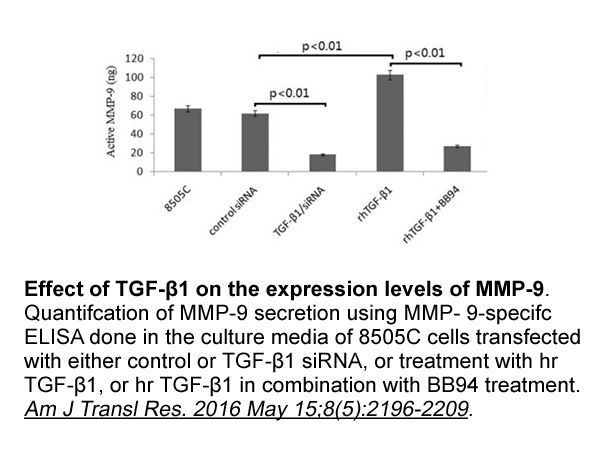
These component origins have been chosen as they Biocytin australia are well-defined options related to developing with components (see for example Morisio and Torchiano (2002); Carney and Long (2000) and Gacek and Arief (2004)). The individual component origins (COTS, OSS, in-house developed co
-
Due to its several industrial applications the recombinant p
2020-01-06
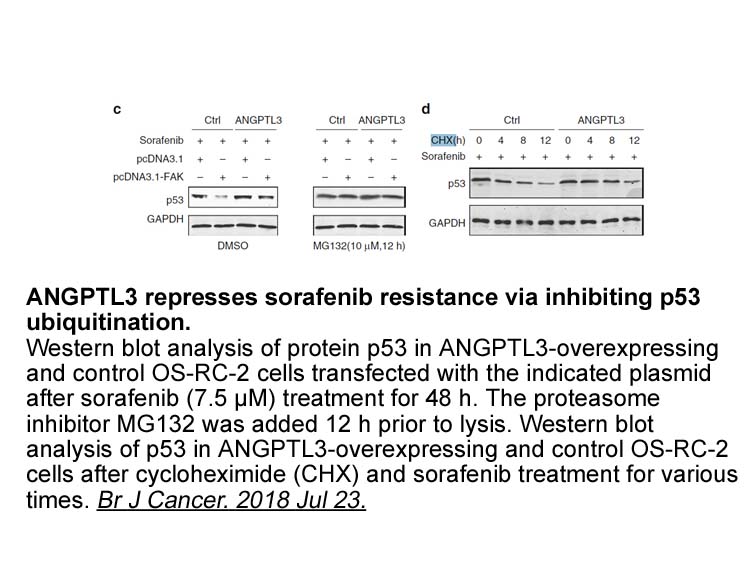
Due to its several industrial applications, the recombinant production of collagen and all its derivatives as gelatins has been pursued for many years in different biological systems as mammalian VU 0155069 australia (Toman et al., 1999), tobacco plants (Ruggiero et al., 2000), silkworms (Tomita et
-
br Materials and methods br Results br Discussion One
2020-01-06
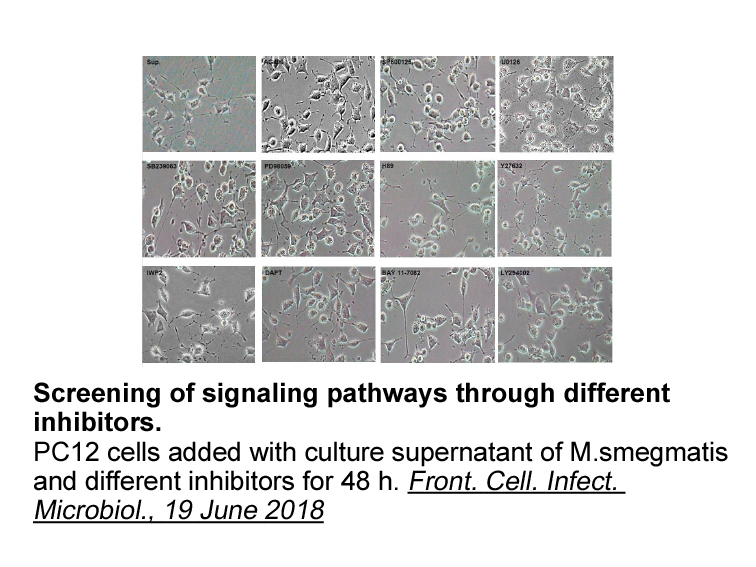
Materials and methods Results Discussion One novel finding in our study is that CK2 inhibition preserved axon function and structure in WM against ischemia. Consistent with these findings, oligodendrocytes, astrocytes, myelin, and egfr inhibitors were found to express CK2α. The robust expr
-
CD 2665 Recently several Pt IV complexes as prodrugs have
2020-01-06
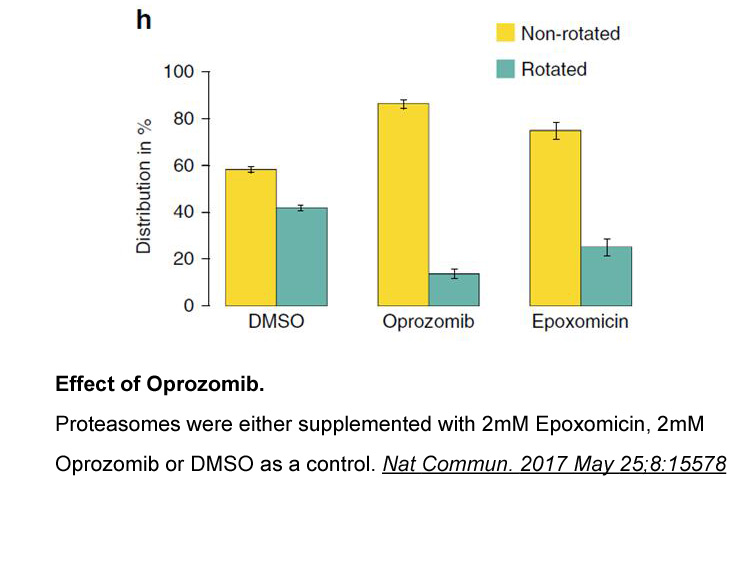
Recently, several Pt(IV) complexes as prodrugs have been designed and studied to enhance anti-tumor efficacy of traditional Pt(II) drugs, improving their cellular uptake and blood-circulation time [26], [27], [28], [29]. Compared with their Pt(II) counterparts, six-coordinated Pt(IV) complexes with
-
Besides the assessment of toxicological effects
2020-01-06
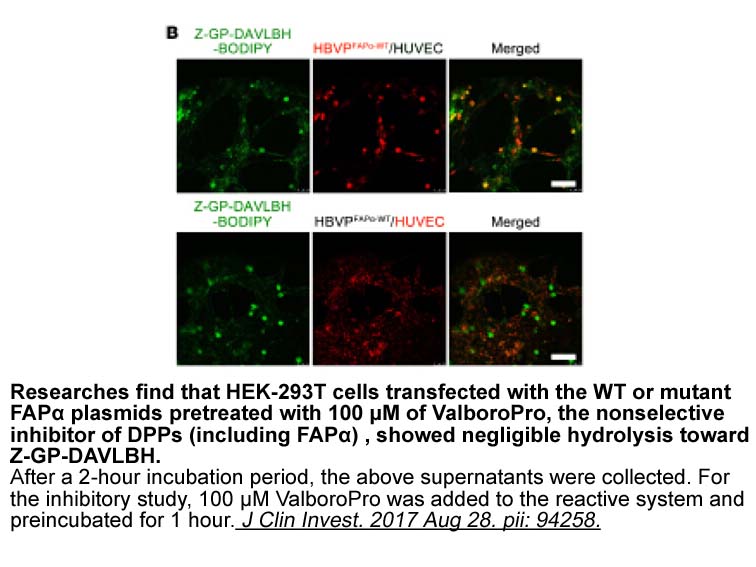
Besides the assessment of toxicological effects immediately after exposure, we also studied the potential recovery of the exposed organisms, through the quantification of ChE activity at specific time intervals after placing organisms in clean test medium. Recovery from chemical challenge, in this c
-
br Materials and methods br Results and discussion br
2020-01-06
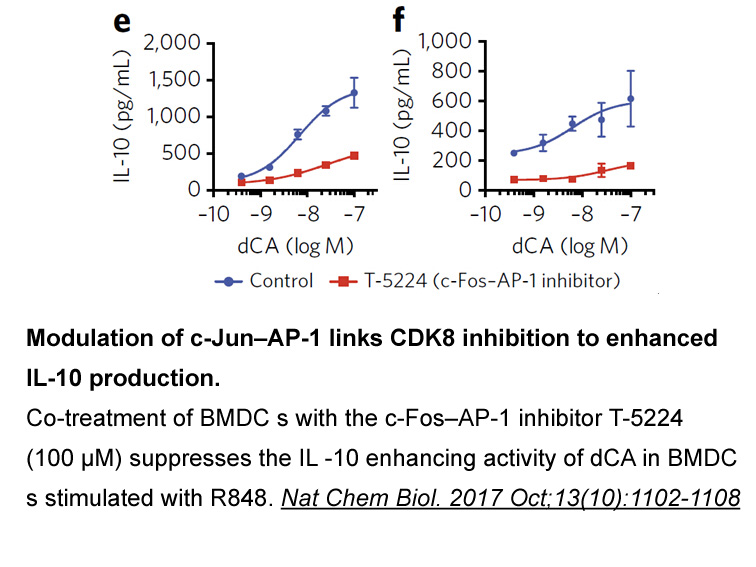
Materials and methods Results and discussion Conclusion The glycyl endopeptidase from papaya latex was partitioned using aqueous two-phase (10%PEG 6000–10% (NH4)2SO4) in combination with ammonium sulphate precipitation (40–60% saturation). The partially purified glycyl endopeptidase showed
-
Using the juxtarenal PPE the induction of juxta
2020-01-06
Using the juxtarenal PPE, the induction of juxta- and suprarenal aneurysms is described herein (Fig. 3A). The infra-/suprarenal part of the CCT251545 analogue synthesis can now be investigated separately, taking into account different embryological backgrounds and varying reactivity to differing sti
15540 records 940/1036 page Previous Next First page 上5页 936937938939940 下5页 Last page